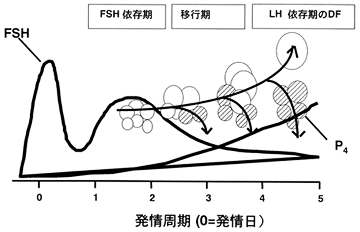
卵胞波(follicular wave)の発現と制御牛の発情周期では,2〜3回の周期性のFSHの増加がみられ,それに伴い連続した卵胞発育の波(卵胞波)の出現がみられる[13, 40, 41].卵胞波では,中型(直径4〜6mm)でエストロジェン活性のある卵胞が2〜5個出現し,そのうち1個の卵胞が選択され(発情周期の2〜3日),持続して発育し,発情周期の4〜5日に優性卵胞となる.優性卵胞が活性を持つ時期には,それ以外の卵胞は閉鎖退行し(図2),FSH濃度は低値となっている[1, 16, 44].同時期に,優性卵胞は発育を続け,卵胞液中のインスリン様成長因子(IGF-1)濃度が増加するとともに,IGF-1結合蛋白-2と-4濃度は低下し,顆粒層細胞のLHレセプタは増加して,エストラジオール分泌を増加させる.そして,LH依存性になり,最終的にはLHパルス頻度により閉鎖するか排卵に至る(図2).黄体期では,3〜4時間ごとのLHパルス頻度で,優性卵胞は優性(dominance)を失い,FSHとLHレセプタおよびエストロジェン産生能力は消失する[16].個々の卵胞には,局所的なフィードバックのループがあり,それはインヒビンやアクチビン,IGF-1やそれぞれの結合蛋白の連続した変化を含み,同様に全身性のゴナドトロピンや成長ホルモン環境下で発達する卵胞の選択および非選択の最終的な運命が決定される[34].インヒビン前躯体の分化過程は,牛の優性卵胞の発育や閉鎖の際に起こる.そこでは総αインヒビン免疫活性の増加と,発情周期の3〜6日に持続的に増加した優性卵胞の大型で単一の形態レベルの継続がみられる[45].優性卵胞のエストロジェン活性の喪失は,総ダイマーインヒビン(二量体インヒビン)と34kDaのダイマー体の増加と関連し,大型分子量体の減少となる[45].
|