![]()
|
ニパウイルス感染症の発生とその考察
白井淳資†(東京農工大学大学院共生科学技術研究院教授)
 1 は じ め に 1 は じ め に1998年にマレーシアで100名を超える死者を出した新しい共通感染症である「ニパウイルス感染症」はあまり関心が持たれていない.狂犬病が我が国で発生していないので警戒されていないのと同様に,遠い国で発生した病気は被害が及ばないと思うのか,マスコミであまり取り上げられたことはない.このニパウイルス感染症は今まで家畜の疾病及び人の病気としては地球上に存在しなかった疾病である.それでは,なぜこのような新興共通感染症が出現したのか.その理由について,私の行ったマレーシアでの研究結果を踏まえ考察してみた.この総説を読んでこのような新しい共通感染症出現の可能性を認識し,今後このような疾病発生を防止する環境作りに努めるようご理解いただけると幸いである. |
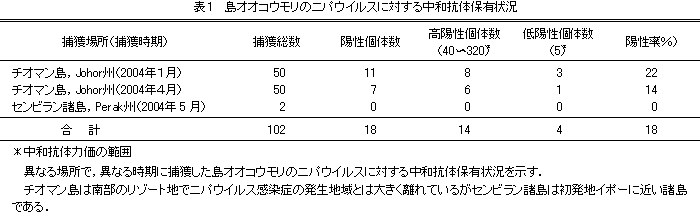
| 2 ニパウイルス感染症の出現 1998年10月より,ペラ州イポーの養豚地域で豚の呼吸器疾病が発生し,1999年の2月まで続いた.本病は,1998年12月頃から発生地のペラ州から感染豚の移動に伴いネグリセンビラン州に広がり,1999年の1月まで続き,1999年1月から3月まで同州の大養豚地域に拡大していったが,それと同時に人の脳炎の発生数も増えていった.1999年の3月にはマレーシアの首都クアラルンプールの近郊セランゴール州にも広がり,集計ではマレーシア国内で265名の患者が確認され,そのうち105名が死亡している.シンガポールでは,マレーシアから輸入した豚を解体する作業に従事していた11名が発症し,そのうち1名が死亡している(図1).本病は豚との接触により人へ感染することが明らかになったため,感染豚の早期摘発淘汰が急務となり,ニパウイルス症状をもとに本病を疑う豚が認められたすべての農場の豚を殺処分した.その後,間接ELISA法と豪州家畜衛生研究所での中和試験による陽性の確認を行い,陽性豚の摘発淘汰を実施した結果,1999年の5月29日に本病の発生は終息した[1]. 1999年3月17日に,ネグリセンビラン州ニパ村出身の脳炎を併発し死亡した人の脳から新種のパラミクソウイルスが分離され,本ウイルスが人及び豚の疾病に関係していることが確認された[2, 3].本病の豚の主たる症状は,呼吸器症状と神経症状である.繁殖豚では流産を起こすものや,神経症状を呈し突然死亡するものが認められた.4〜6週齢の豚では呼吸器症状を示すものが多く,犬の吠え声のような大きな咳をするので,Barking illとも呼ばれていた.豚がニパウイルスに感染したかどうかは,39.9℃以上の発熱と激しい発咳,腹式呼吸もしくは開口呼吸などを伴う呼吸器症状が認められ,神経性の振戦や痙攣,後躯麻痺,痙攣麻痺,また豚が追い立てられたり,駆けだしたときに認められる歩行不正などの神経症状のうちどれか一つもしくはそれ以上を示すことにより診断した.養豚場で繁殖雌豚や雄豚の突然死が見られることなども本病の特徴であった.豚において本病の死亡率は5%以下と低いものの,感染率は80%以上と高く,症状を示さない豚が多く存在した[1]. ニパウイルス感染症発生後の抗体調査により豚以外の感染動物は,犬での感染率が高く,続いて馬,山羊,猫や齧歯類まで感染が認められている.犬は本病に容易に感染するため,発生周辺地域の犬は捕獲され殺処分された.しかしその後の調査では犬間の感染は認められておらず,馬,猫及び山羊などは豚から感染したもので,これらの動物間での感染は認められていない[3].豚以外の動物から人への感染は確認されなかった.広範な野生動物の抗体調査から[4],本ウイルスの自然宿主としてオオコウモリにニパウイルスのELISA抗体が確認されていたため,2002年より国際協力機構(Japan Cooperation Agency : JICA)のプロジェクトとして3年間マレーシア国立獣医学研究所と共同研究を行い,どのようにしてオオコウモリから豚に感染したか疫学調査を行った.以前の調査でELISA抗体の陽性が認められた,マレーシアのどこでもみられる小型の果樹食性(Cynopterus brachyotis 及びEonycteris spelaea )オオコウモリは木陰(写真1)や洞窟を営巣場所とし,夕暮れ農家の庭先の果樹に多く飛来するので,簡単に捕獲することができた.全部で500頭あまりのCynopterus brachyotis 及びEonycteris spelaea を調べたが,ニパウイルスの中和抗体は検出されず,ニパウイルスも分離されなかった.しかし,マレーシア周辺の小島の海岸に生息し営巣する島オオコウモリ(Pteropus hypomeranus)(写真2,3)には平均18%の割合でニパウイルスに対する中和抗体が存在し(表1),ニパウイルスも分離されている[5].しかし,島オオコウモリの飛行距離は短く,生息している島から出ることはないのでニパウイルス感染症発生原因とは考えにくい.そこで,マレー半島の河口付近のマングローブジャングルに生息し営巣する大型のオオコウモリであるマラヤオオコウモリ(Pteropus vampyrus)(写真4,5)を捕獲しニパウイルスに対する中和抗体保有率を調べたところ,なんと平均63%もの高い割合を示した(表2).ニパウイルスの分離には成功しなかったものの,マラヤオオコウモリの気管拭い液からは,リアルタイムRT-PCRでニパウイルスの遺伝子を検出した.これらの結果から,マレーシアで発生したニパウイルス感染症は,このマラヤオオコウモリから養豚場の豚にウイルスが感染し,豚の間で広がってゆき,養豚に従事し豚に接触した人にこの病気が感染し,新しい共通感染症が出現したものと考えられた.しかし,マラヤオオコウモリはずっと昔からジャングルに生息し,今まで問題を起こすことなく悠久の時を過ごしてきた.この時期になってなぜマラヤオオコウモリから新興共通感染症が広がるようになったのか考察してみた.その理由としては,1990年よりシンガポールが完全に畜産業を行わないことを決めたためにシンガポールへの豚の輸出件数が増え,この時期よりマレーシアにおける養豚産業が急激に成長してきた.それに伴う養豚場拡充のためオオコウモリの餌場であるジャングルを切り開き,どんどん養豚場を建設していったのである.また,マレーシアはイスラム教国家であることもあり,養豚を営むための制限は厳しく,養豚場の周囲には必ず空所を設けなければならない[6].養豚主はこの空所に果樹を植えて,養豚業と果樹園の両方から収益を得ようとした.また,ニパウイルス感染症が発生した1997年から1998年頃には異常気象の影響で雨が降らず,ジャングルの果樹が枯渇した[7].このような条件が重なり,普段はジャングルで静かに暮らしているマラヤオオコウモリが養豚場周辺の果樹園に降りてきて作餌するようになった.オオコウモリは飛行する動物であるので,体重の増加を抑えるため固形物を摂食せず,液体成分だけ吸飲し,残りカスはチューインガムの噛みカスのようにはき出す.この噛みカスを豚が摂食し,オオコウモリが保有していたニパウイルスが豚に感染し,豚の間で蔓延したのだろうと考えている.豚の症状は呼吸気症状が主であったため,豚から豚へ次々と感染して行き,豚の世話をしていた人にも空気伝搬により感染していったのである. |
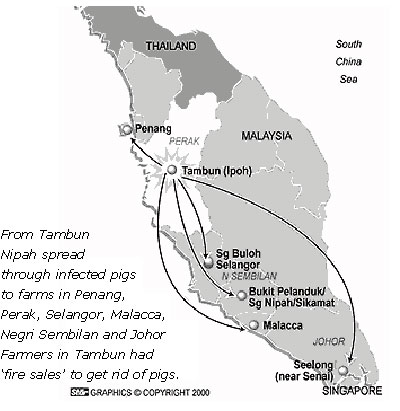 (2000年のマレーシア農業省ニパウイルス感染症発生報告書) 図1 イポーのTambun地区から感染豚の異動により,ペナン,セランゴール,マラッカ,ネグリセンビラン及びジョホール州の養豚地域に広がった.Tambun地区の養豚業者は感染豚の激安売り(Fire sale)で病気を広げた. |
 写真1 公園や民家近くの木陰に営巣する小型フルーツバット(Cynopterus brachyotis).マレーシアでは各地で頻繁にみられる. |
 写真2 島オオコウモリの営巣場所 島オオコウモリ(island flying fox : Pteropus hypomeranus)は海岸に植える椰子の木やその他の木に営巣する.写真中央の民家の後部の木に鈴なりにぶら下がる島オオコウモリ(○枠内) |
 写真3 島オオコウモリ(Ptropus hypomeranus) 島オオコウモリの体長は約17cmで翼長は1mほどである.体重は約500gで毛色は焦げ茶色である.飛行距離は短く生息する島より外に出ることはない. |
 写真4 マラヤオオコウモリの営巣場所 マラヤオオコウモリ(Malayan flying fox : Pteropus vampyrus)は河口付近のマングローブジャングル内の木に営巣する(○枠内).木の下の地面は水没し,人は容易に近づけない. |
 写真5 マラヤオオコウモリ(Pteropus vampyrus) マラヤオオコウモリの体長は約25cmで翼長は1.2mに及ぶ.体重は約1kgで島オオコウモリより一回り大きく,毛色は黒で背部の一部のみ焦げ茶色である.飛行距離は長く国境を越えタイ南部およびインドネシアジャワ島まで飛行し,最大250kmに及ぶと言われている. |